生物化学_Lipid Biosynthesis
- Lipid Biosynthesis
- 发生地点
- Eukaryotic Lipids are Made at a Variety of Sites
- Occurs mainly in liver and adipocytes (mammals)
- Most lipid biosynthesis in eukaryotic cells occurs in the ER.
- Enzymes of lipid synthesisare membrane-bound with active sites facing the cytosol.
- Other lipid synthesis locations include - plasma membrane, mitochondria, lysosomes and peroxisomes
- fatty acid synthesis vs degradation
合成与消耗反应的区别- Intermediates in fatty acid synthesisare linked covalently to the sulfhydryl 巯基 groups of special proteins, the acyl carrier proteins (ACPs). Incontrast, fatty acid breakdown intermediates are bound to the-SH group of coenzyme A.
- Fatty acid synthesis occurs in the cytosol, whereas fatty acid degradation takes place in mitochondria.
- In animals, the enzymes of fatty acid synthesis are components of one long polypeptide chain, the fatty acid synthase, whereas no similar association exists for the degradative enzymes.
- The coenzyme for the oxidation–reduction reactions of fatty acid synthesis is NADP+/ NADPH, whereas degradation involves the NAD+/ NADH couple.
- 原料&能量
- Acetyl-CoA
- Amino acid degradation produces cytosolic acetyl-CoA.
normally insufficient for fatty acidbiosynthesis
- Amino acid degradation produces cytosolic acetyl-CoA.
- Fatty acid oxidation produces mitochondrial acetyl-CoA.
cannot cross the mitochondrial membrane
- Fatty acid oxidation produces mitochondrial acetyl-CoA.
- Glycolysis yields cytosolic pyruvate, which is converted to acetyl-CoA by pyruvate dehydrogenase
cannot cross the mitochondrial membrane
- Glycolysis yields cytosolic pyruvate, which is converted to acetyl-CoA by pyruvate dehydrogenase
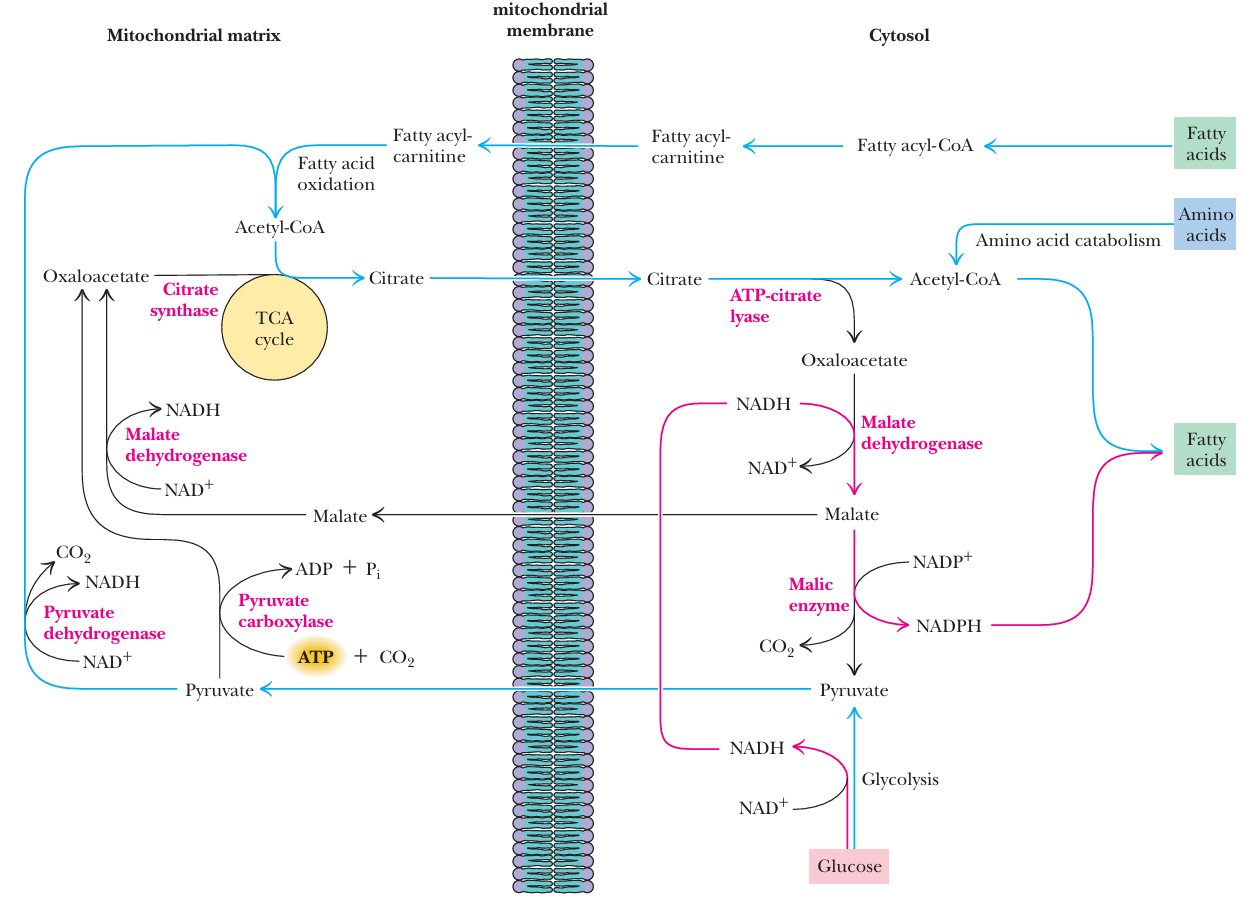
- 2/3来源的acetyl-CoA不能直接跨线粒体膜, 因此需要 acetyl-CoA + oxaloacetate →(citrate synthase) citrate → 跨膜 → citrate →(ATP-citrate lyase) acetyl-CoA + oxaloacetate
- Reducing Power - NADPH
- NADPH can be produced in the pentose phosphate pathway as well as by malic enzyme.
- Reducing equivalents (electrons) derived from glycolysis in the form of NADH can be transformed into NADPH by the combined action of malate dehydrogenase and malic enzyme.
NADH不能用于fatty acid合成, 借助合成再分解malate变成NADPH
- 蓝线代表carbon, 红线代表electrons
- Acetyl-CoA
- Acetyl-CoA → Malonyl-CoA
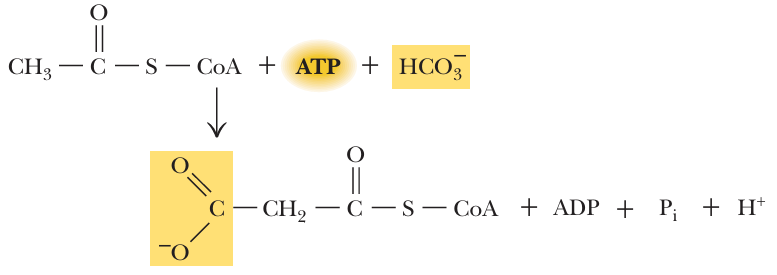
- The carboxylation of acetyl-CoA to form malonyl-CoA is essentially irreversible and is the committed step in the synthesis of fatty acids.
a “committed step” is an effectively irreversible enzymatic reaction that occurs early in a metabolic pathway, once this step happens, the molecule is “committed” to continuing down that specific pathway to the end product - The reaction is catalyzed by acetyl-CoA carboxylase,which contains a biotin维生素B7 prosthetic group辅基
- This carboxylase is the only enzyme of fatty acid synthesis in animals that is not part of the multienzyme complex called fatty acid synthase
- The acetyl-CoA carboxylase reaction produces malonyl-CoA for fatty acid synthesis
- The carboxylation of acetyl-CoA to form malonyl-CoA is essentially irreversible and is the committed step in the synthesis of fatty acids.
- Acyl Carrier Proteins (ACP) Carry the Intermediates in Fatty Acid Synthesis
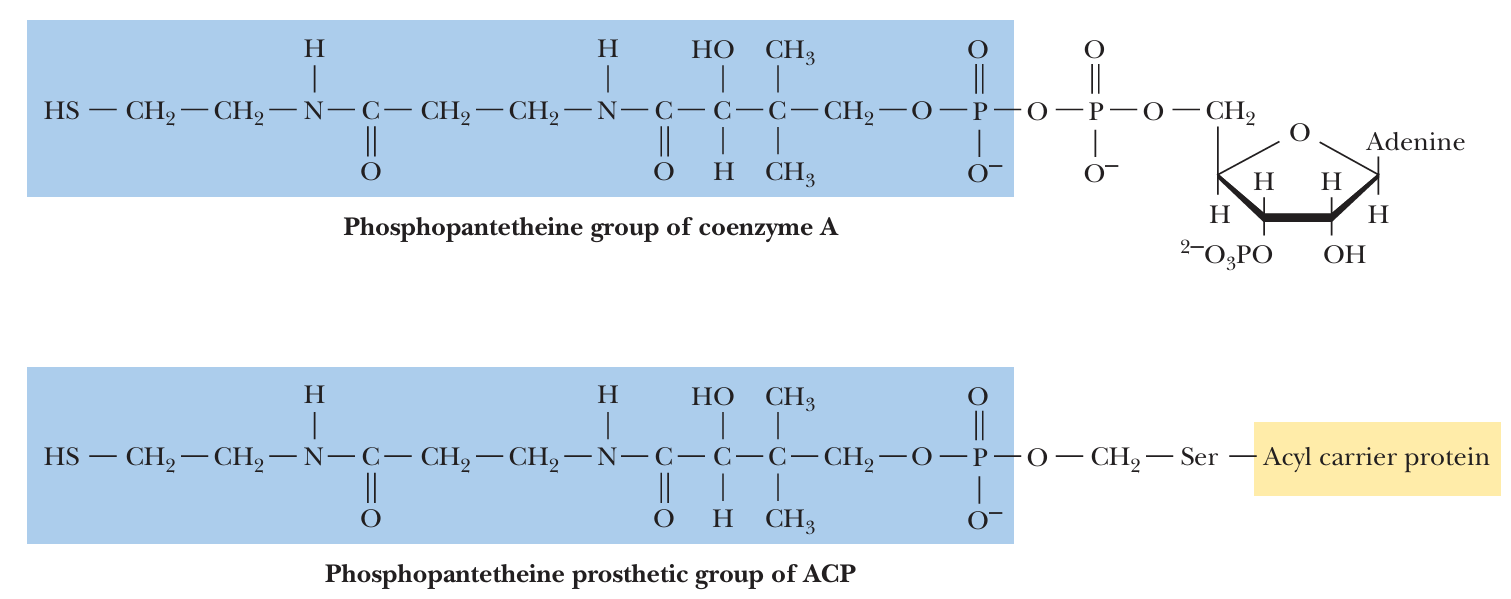
- The basic building blocks of fatty acid synthesis are acetyl and malonyl groups, but they are first passed to Acyl Carrier Proteins (ACP)
- ACP consists (in E. coli) of a single polypeptide chain of 77 residues to which is attached (on a serine residue) a phosphopantetheine group磷酸泛酰巯基乙胺基团, the same group that forms the “business end” of coenzyme A.
fatty acid 就是通过蓝色这个p巴拉巴拉集团连接到CoA/ACP上的 - Thus, ACP is a somewhat larger version of coenzyme A, specialized for use in fatty acid biosynthesis.
- ACP 相当于合成 fatty acid 的专用 CoA
- In Some Organisms, Fatty Acid Synthesis Takes Place in Multienzyme Complexes
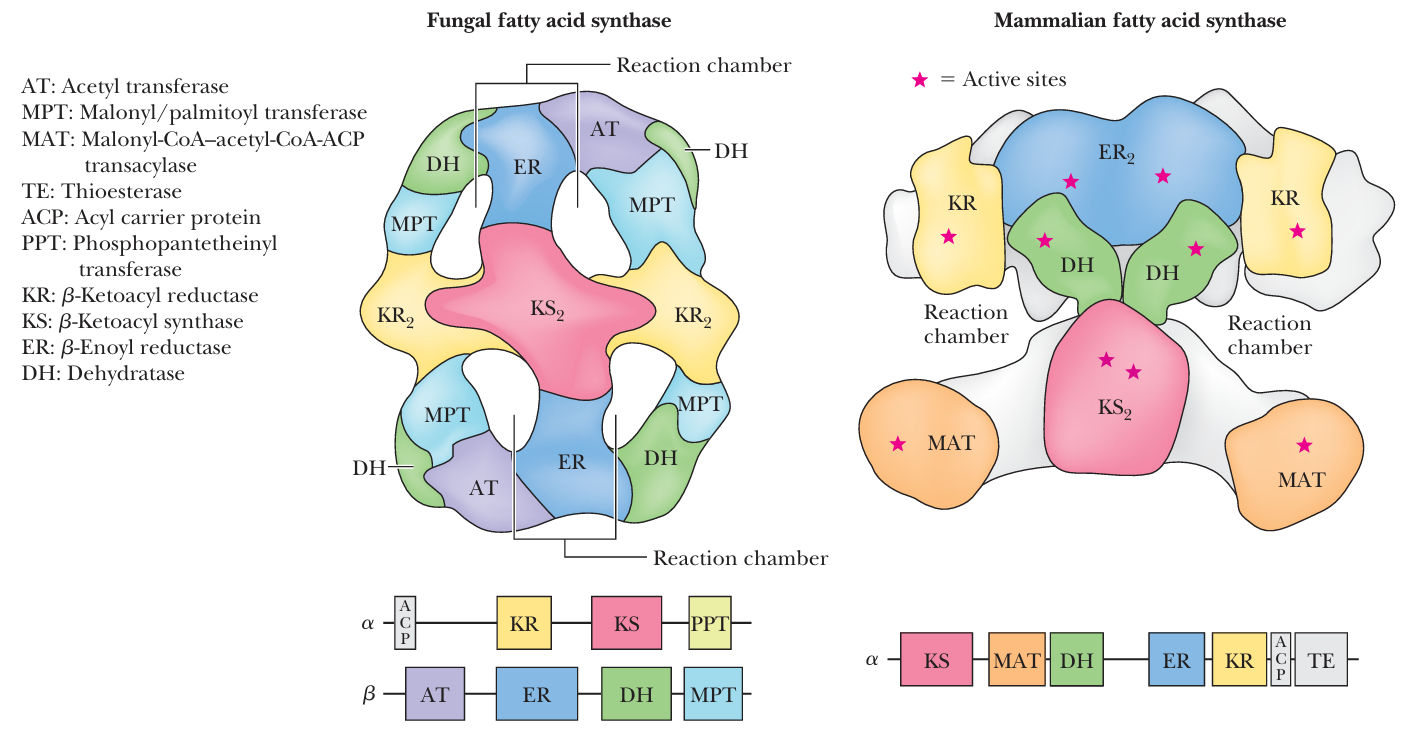
- fatty acyl synthase I (FAS I)
- FAS in mammals on homodimeric FAS I, each 270-kD polypeptide of which contains all reaction centers required to produce a fatty acid
- FAS in lower eukaryotes, such as yeast and fungi, the enzymatic activities of FAS are distributed on two multifunctional peptide chains
- 图
- In both the fungal and the mammalian FAS structures, the close association of enzymic domains within one large complex permits efficient transfer of intermediates from one active site to the next.
- fatty acyl synthase II (FAS II)
- In plants, most bacteria, and parasites寄生虫
- the enzymes of fatty acid synthesis are separated and independent
- this collection of enzymes is referred to as FAS II
- fatty acyl synthase I (FAS I)
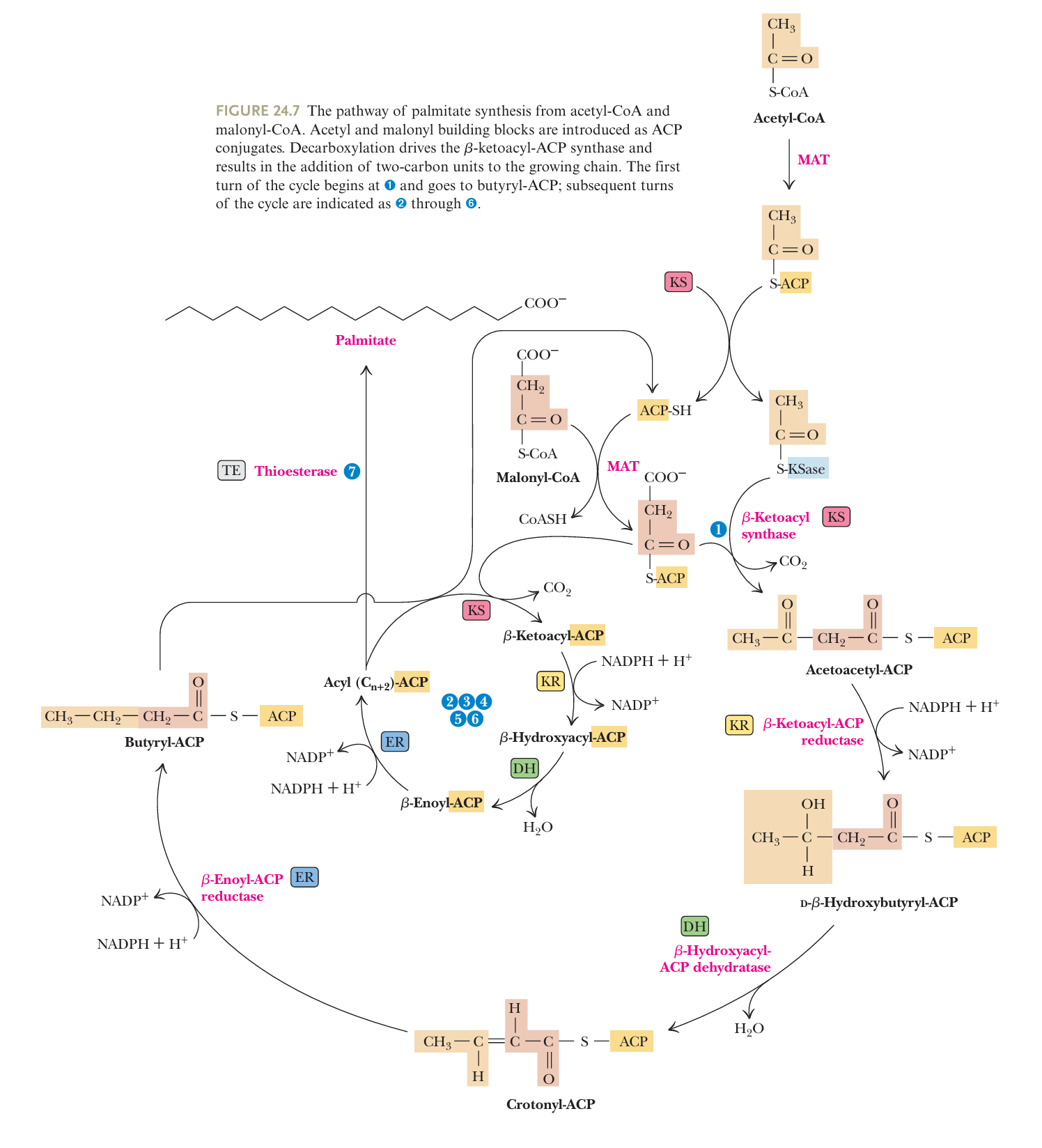
- C16 fatty acid synthesis reaction 核心反应
- 外面是第一圈, 中间是后续循环, 原理一样, 底物长度逐渐增加
- 第一圈把 Acetyl-ACP / Malonyl-ACP 接一块, 后续在接一块的butyryl上面循环加2个C, 第一圈和后续循环的原理是一样的, 除了底物C每次加2
- Acetyl-CoA / Malonyl-CoA 上 ACP
起手
- initiate elongation cycle
- 酶 - malonyl-CoA–acetyl-CoA-ACP transacylase (MAT)
- Acetyl-CoA / Malonyl-CoA 上 ACP
- KS操作
合体长链/acetyl 与 malonyl-ACP
- The β-ketoacyl-ACP synthase (KS) catalyzes the decarboxylative condensation of the acyl group with malonyl-ACP to produce a beta-ketoacyl酮脂酰-ACP intermediate (acetoacetyl-ACP in the first cycle)
- hydrolysis of ATP drove the carboxylation of acetyl-CoA to form malonyl-ACP, so indirectly ATP is responsible for the condensation reaction to form acetoacetyl-ACP, and Malonyl-CoA can be viewed as a form of stored energy for driving fatty acid synthesis
能量流动 ATP → malonyl-ACP → fatty acid
- KS操作
- Reduction of the β-Carbonyl Group
一系列操作删除C=O, 饱和碳链
- ① reduction of the β-carbonylgroup by β-ketoacyl-ACP reductase (KR) to form a β-alcohol
C=O → C-OH - ② dehydration by β-hydroxyacyl-ACP dehydratase (DH)
C-OH → C=C - ③ reduction by 2,3-trans-enoyl-ACP reductase (ER)
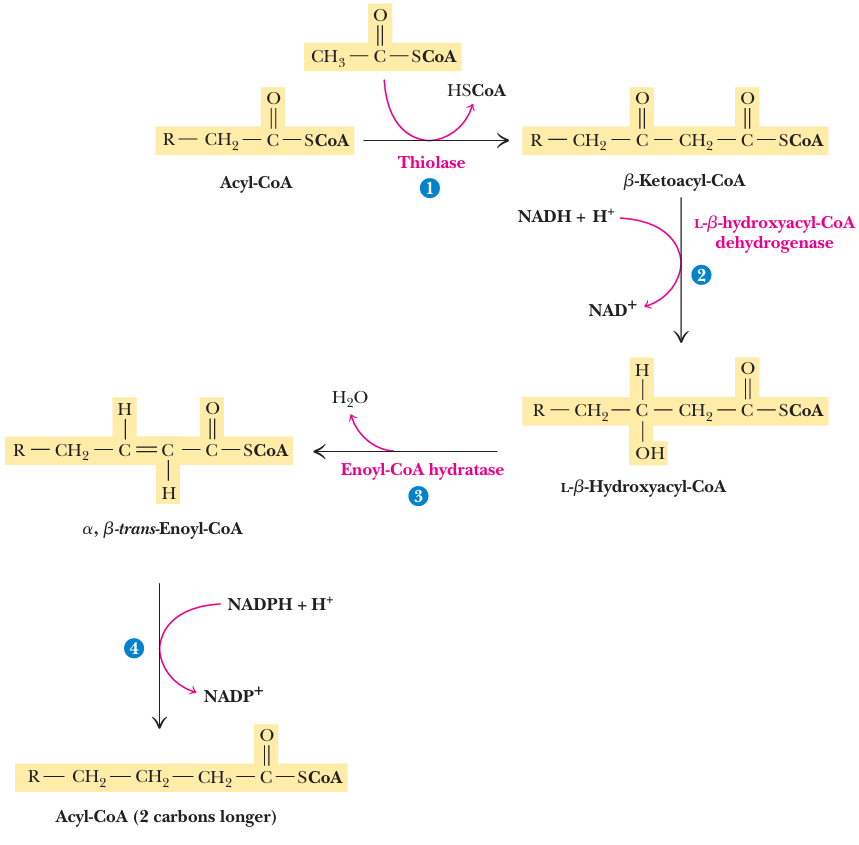
C=C → C-C - 与fatty acid degradation in reverse类似, 但有区别
- different enzymes are involved
- the alcohol formed in biosynthesis has the D-configuration rather than the L-form seen in catabolism
- the reducing coenzyme is NADPH, whereas NAD+ and FAD+ are the oxidants in the catabolic pathway
- Reduction of the β-Carbonyl Group
- end
- This cycle continues with the net addition of a two carbon unitin each turn until the chain is 16 carbons long.
- The KS cannot accommodate larger substrates, so the reaction cycle ends with a 16-carbon chain
- 外面是第一圈, 中间是后续循环, 原理一样, 底物长度逐渐增加
- C16 fatty acid elongation
- Longer chains are made through special elongation reactions, which occur both in the mitochondria and at the surface of the ER.
- The ER reactions
- are similar to former - addition of two-carbon units at the carboxyl end of the chain by the oxidative decarboxylations of malonyl-CoA
- this decarboxylation provides the thermodynamic driving force for the condensation reaction
- The mitochondrial reactions
- involve addition of acetyl units
- These reactions are essentially a reversal of fatty acid oxidation, with the exception that NADPH is utilized in the saturation of the double bond, instead of FADH2
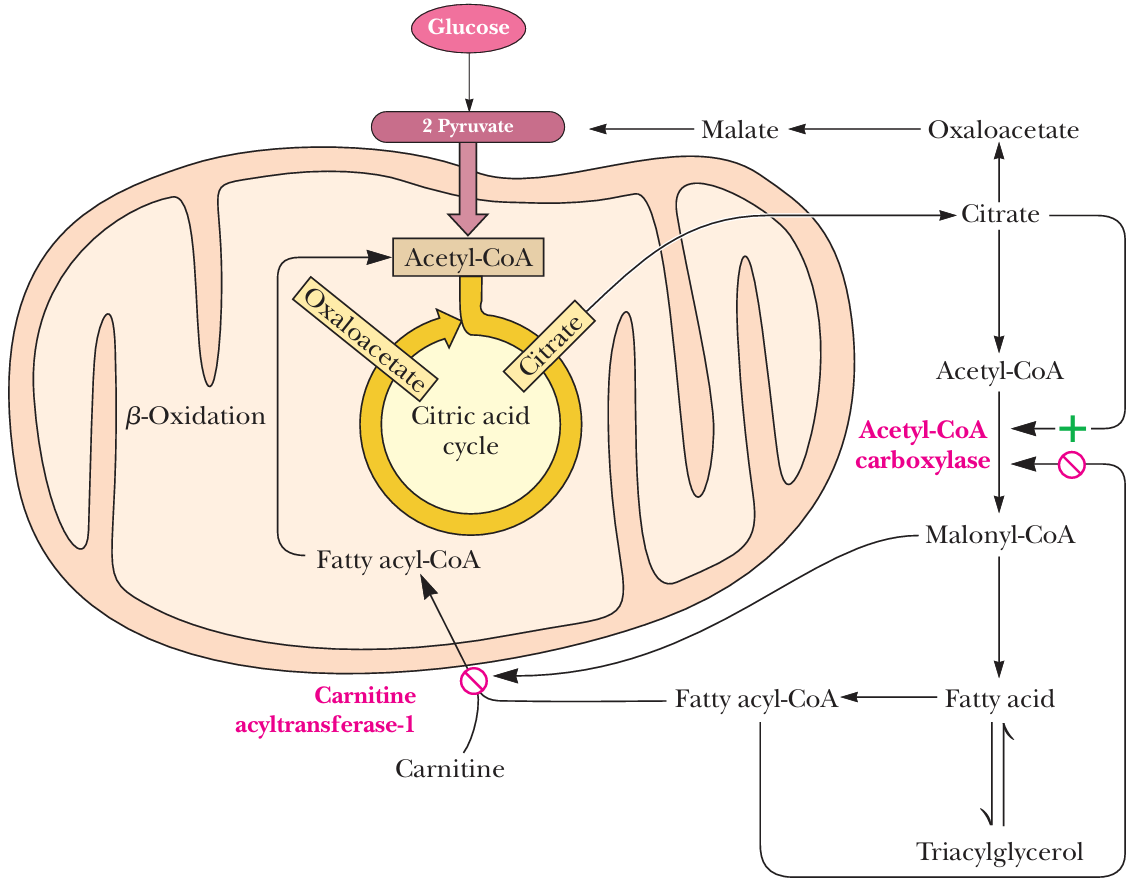
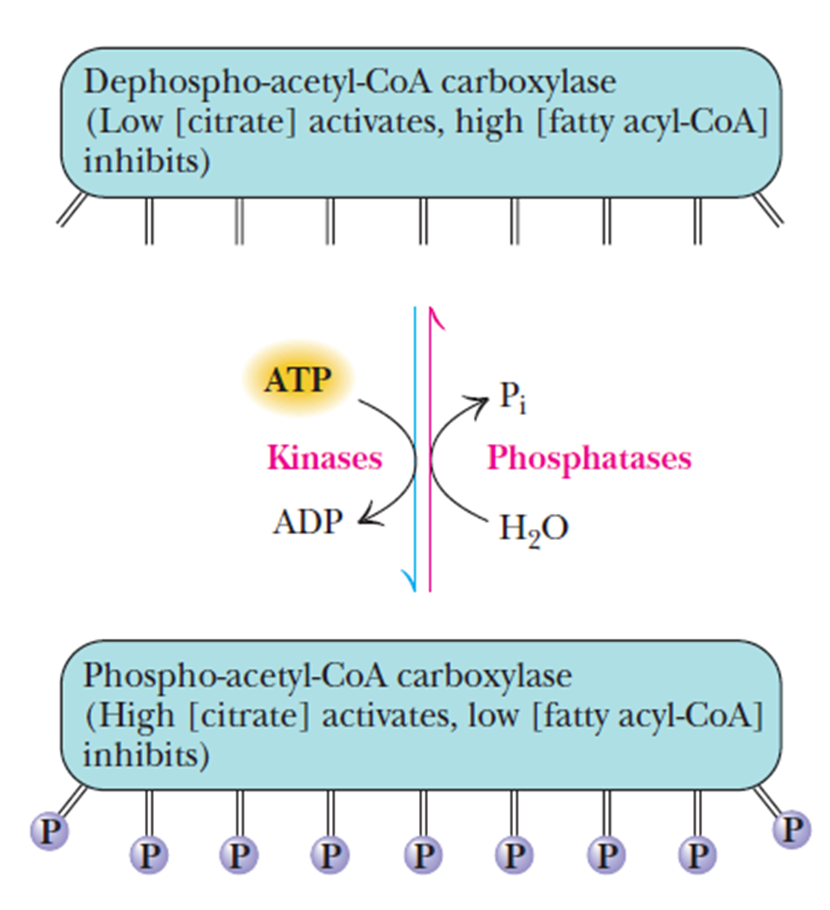
- Regulation of fatty acid synthesis and oxidation
- 产物对两个方向的负反馈调节
- 图
- Malonyl-CoA
- inhibiting the carnitine acyltransferase
- inhibits the uptake of fatty acylcarnitine 脂肪酰肉碱 (and thusfatty acid oxidation) by mitochondria.
- 在生产fatty acid过程中不让线粒体进行β-oxidation
- citrate
- levels rising (which reflect anabundance of acetyl-CoA) signal the initiation of fattyacid synthesis
- an important allosteric activator of acetyl-CoA carboxylase
- fatty acyl-CoA
- when levels rise, fatty acid synthesisis inhibited and fatty acid oxidation activity increases
- an important allosteric inhibitor of acetyl-CoA carboxylase
- The degree of inhibition is proportional to the chain length of the fatty acyl-CoA
碳链越长, 别构抑制作用越强
- 图
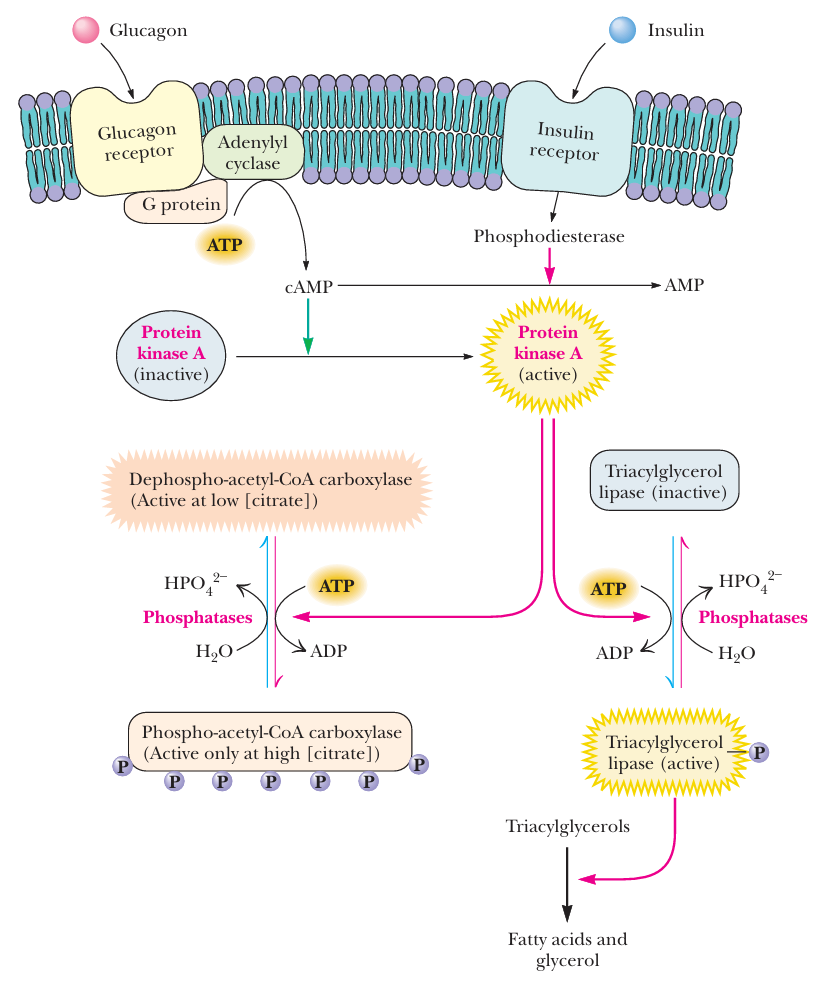
- Hormonal Signals
- 图
- hormone
- glucagon 胰高血糖素
- G protein → cAMP → protein kinase A → phosphate ACC / phosphate lipase
- insulin 胰岛素
- receptor → cAMP ↓ 抵制胰高血糖素
- epinephrine 肾上腺素
- activate lipase
- glucagon 胰高血糖素
- ACC
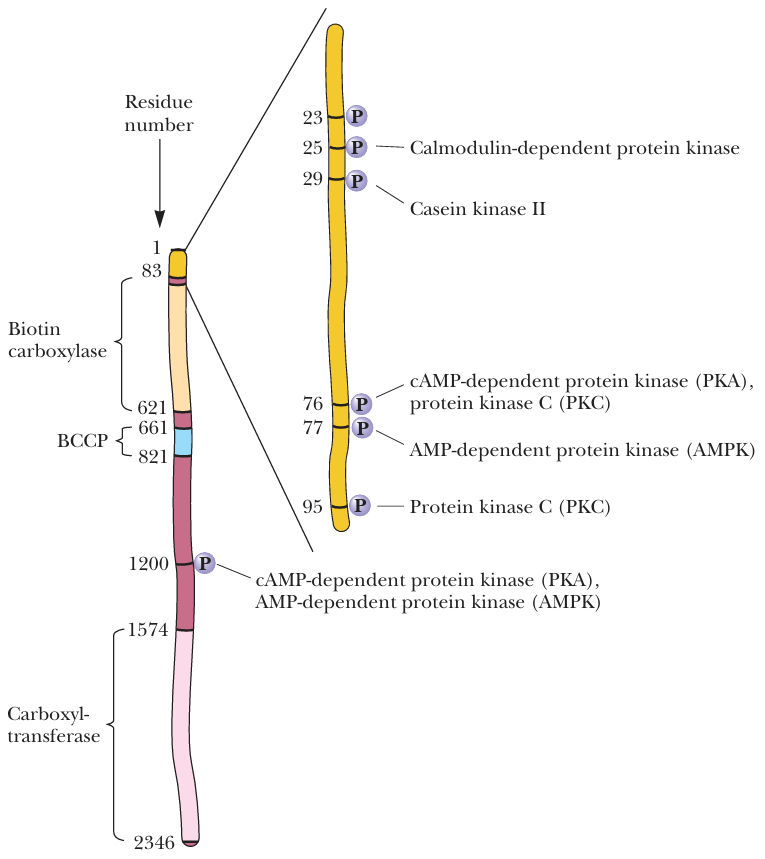
- 结构图, Phosphorylation at Ser1200 is primarily responsible for decreasing the affinity for citrate
- 2种状态
- 1200没P
- 不会降低对citrate的affinity
- 高浓度脂肪才会抑制
- 这种状态利于合成脂肪
- 1200有P
- 会降低对citrate的affinity
- 低浓度脂肪就会抑制
- 这种状态不利于合成脂肪
- 图, 由于ACC单体串成一串, 示意图中每个P代表不同单体1200位置
- 1200没P
- glucagon可以将它切换到有P状态, 抑制脂肪合成
- PP2A可以将它切换到无P状态, 促进脂肪合成
- 这是五碳糖途径促进脂肪合成的一部分体现, 五碳糖途径可以促进脂肪合成 → Regulation of fatty acid synthesis and oxidation
- 结构图, Phosphorylation at Ser1200 is primarily responsible for decreasing the affinity for citrate
- lipase
- glucagon / epinephrine 都可以激活它
- lipase 将 triacylglycerol 分解为 fatty acid 与 glycerol
- 分解出来的 fatty acid 进入线粒体进行 β-oxidation, 释放能量
- 这个酶促进脂肪分解
- 图
- 产物对两个方向的负反馈调节
- lipid transport
- When most lipids circulate in the body, they do so in the form of lipoprotein complexes.
- 当大多数脂质在体内循环时,它们是以脂蛋白复合物的形式进行的
- Simple, unesterified fatty acids are only bound to serum albumin and other proteins in blood plasma, but phospholipids, triacylglycerols, cholesterol, and cholesterol esters are all transported in the form of lipoproteins.
- 简单的未酯化脂肪酸仅与血浆中的血清白蛋白和其他蛋白质结合, 而磷脂、甘油三酯、胆固醇和胆固醇酯则全部以脂蛋白的形式进行运输。
- At various sites in the body, lipoproteins interact with specific receptors and enzymes that transfer or modify their lipid cargoes.
- 在身体的不同部位,脂蛋白会与特定的受体和酶发生相互作用,从而转移或修饰它们所携带的脂质载荷
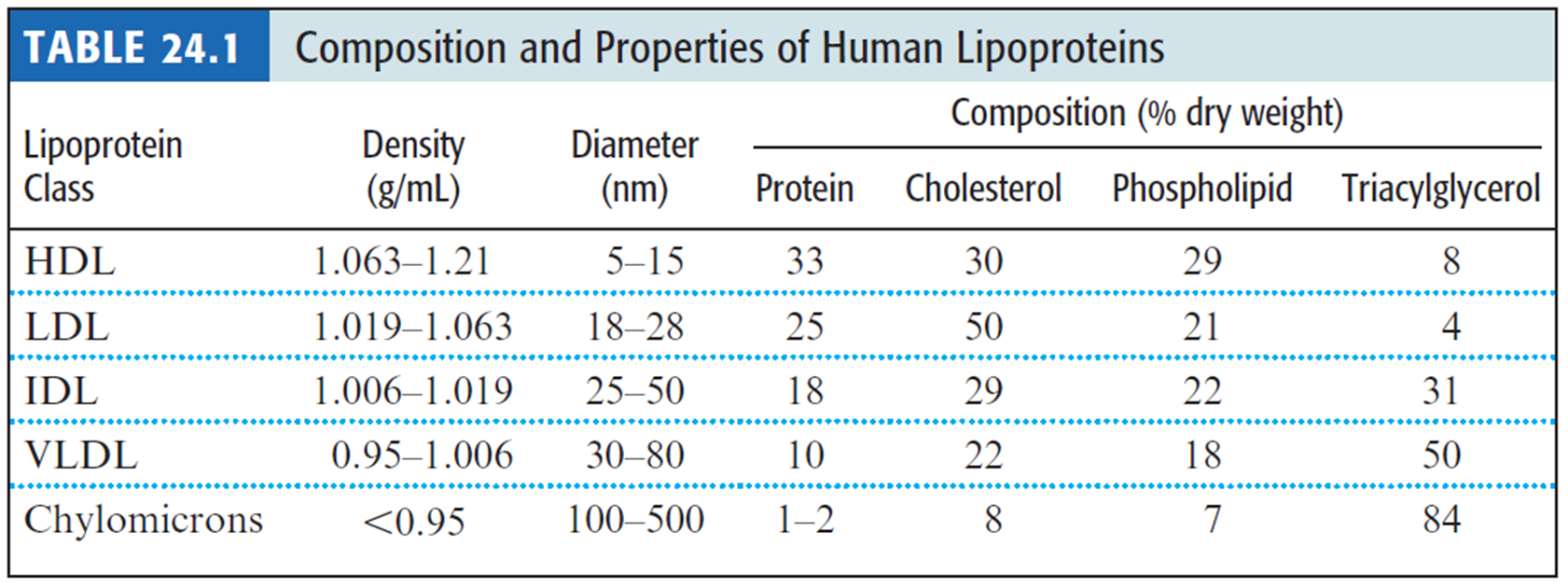
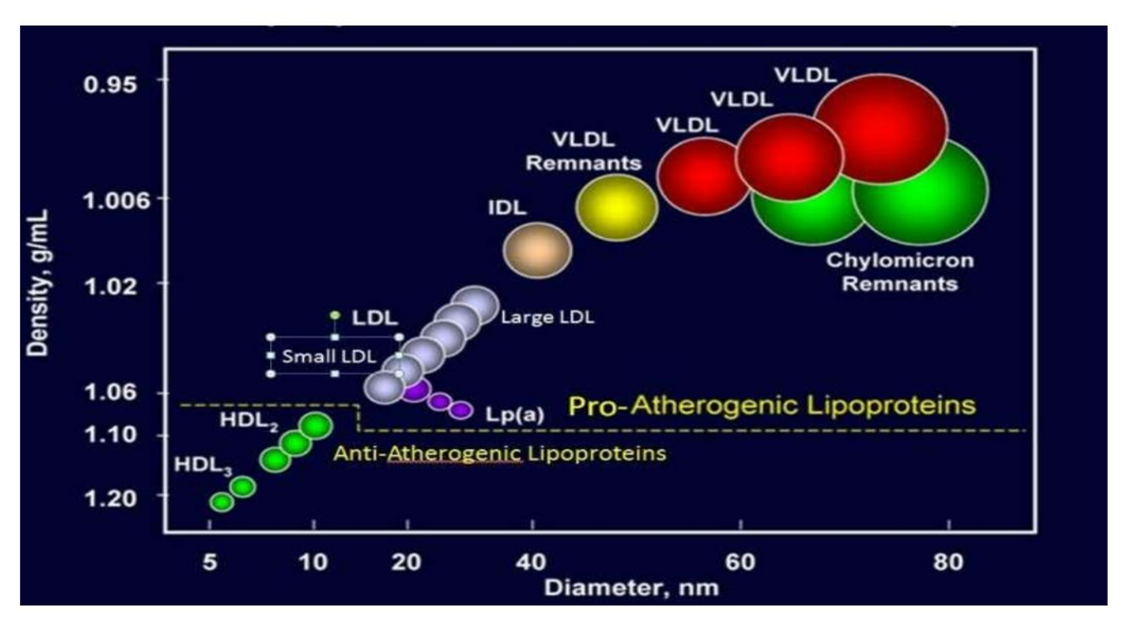
- Lipoproteins were classified according to their densities.
- 脂蛋白是根据它们的密度进行分类的
- The densities are related to the relative amounts of lipid and protein in the complexes.
- 密度与复合物中脂质和蛋白质的相对含量有关
- Because most proteins have densities of about 1.3 to 1.4 g/mL, and lipid aggregates usually possess densities of about 0.8 g/mL, the more protein and the less lipid in a complex, the denser the lipoprotein.
- 由于大多数蛋白质的密度约为 1.3 至 1.4 g/mL,而脂质聚集体的密度通常约为 0.8 g/mL,因此复合物中蛋白质越多、脂质越少,脂蛋白的密度就越大
- there are high-density lipoproteins (HDLs), low-density lipoproteins(LDLs), intermediate-density lipoproteins (IDLs), very-low-density lipoproteins(VLDLs), and also chylomicrons 乳糜微粒
- Pro-Atherogenic 促动脉粥样硬化 lipoproteins, Anti-atherogneic 抗动脉粥样硬化 lipoproteins
- Chylomicrons
- the largest lipoproteins.
- They deliver TGs甘油三酯 from the intestine (via lymph and blood) to tissues (muscle for energy, adipose for storage).
- They are present in blood only after feeding.
- Cholesterol-rich chylomicron remnants 残留物deliver cholesterol to the liver.
富含胆固醇的乳糜微粒残留物(chylomicron remnants)将胆固醇运送到肝脏
All articles in this blog are licensed under CC BY-NC-SA 4.0 unless stating additionally.
